

The epigenome is the broad array of chemical marks that decorate DNA and its protein scaffold. These marks act like a chemical notation, telling the cell which genes to express and which to keep silent. As such, the epigenome helps to explain how cells with identical DNA can develop into the multitude of specialized types that make up different tissues. The marks help cells in the heart, for example, maintain their identity and not turn into neurons or fat cells. Misplaced epigenetic marks are often found in cancerous cells.
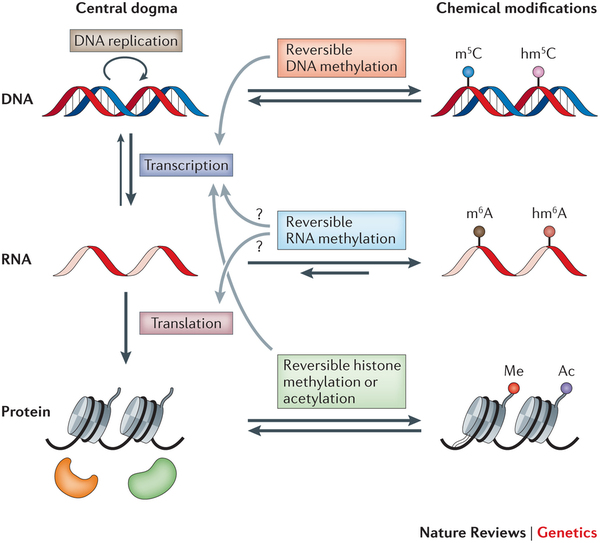
Most epigenetic research focused on the tags associated with DNA and the histone proteins that it wraps around. But more than 100 different types of chemical mark had been identified on RNA, and nobody knew what they did.
In 2009, researchers began hunting for reversible marks on RNA and the proteins that erase them.
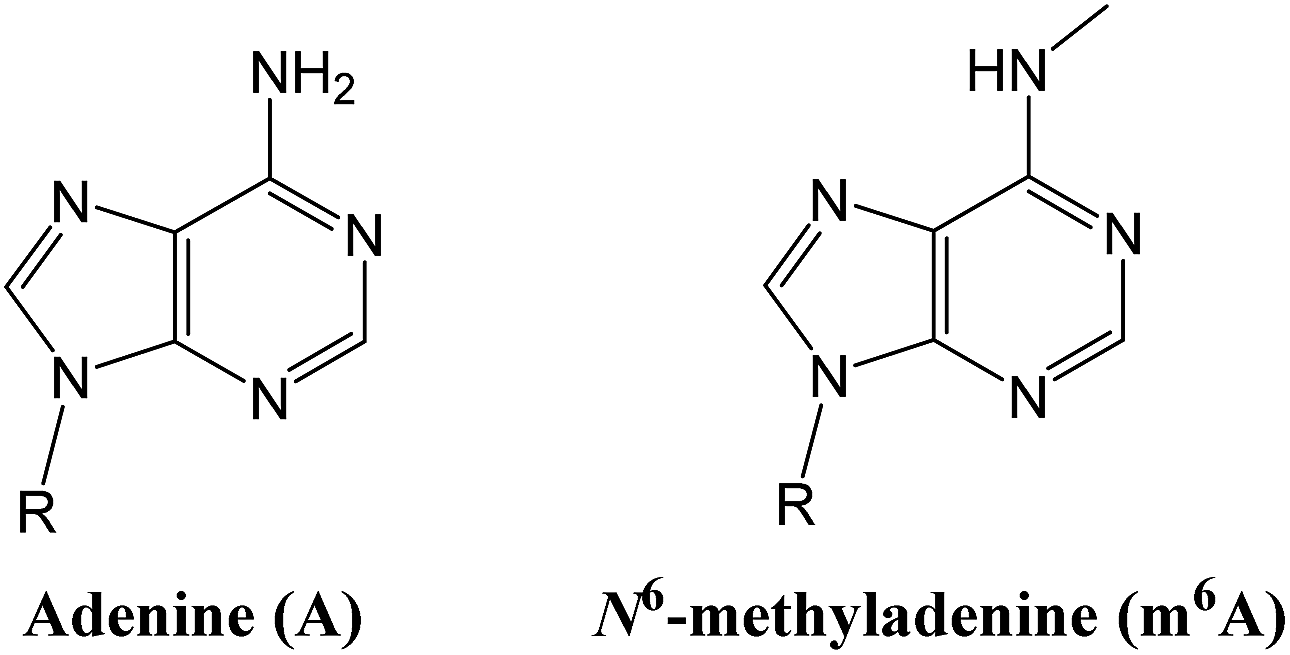
Nine years later, such research has given birth to an ‘ome of its own, the epitranscriptome. Researchers have shown that a methyl group attached to adenine, one of the four bases in RNA, has crucial roles in cell differentiation, and may contribute to cancer, obesity and more. In 2015, He’s lab and two other teams uncovered the same chemical mark on adenine bases in DNA (methyl marks had previously been found only on cytosine), suggesting that the epigenome may be even richer than previously imagined.
The governing rule of molecular biology — the central dogma — holds that information flows from DNA to messenger RNA to protein. Many scientists therefore viewed mRNA as little more than a courier, carrying the genetic information encoded in a cell’s nucleus to the protein factories in the cytoplasm. That’s one reason why few researchers paid much attention to the modifications made to mRNA.
In 2010 the team decided to test FTO’s activity on m6A — the methylated adenine. The mark disappeared. The team had shown for the first time that RNA methylation was reversible, just like the marks found on DNA and histones. To He, it seemed like proof of an RNA-based system of gene regulation.

Evidence mounts
He’s group wasn’t the only one thinking about m6A. In 2012, two teams of researchers independently published the first maps of where m6A appears. The studies revealed more than 12,000 methylated sites on mRNAs originating from about 7,000 genes.
One m6A reader, for example, makes mRNA degrade faster by shuttling it to decay sites in the cell. Another m6A reader promotes protein production by shepherding methylated RNA to the ribosome.
Whether m6A directs a cell to produce a protein or destroy a transcript depends on the location of the mark and on the reader that binds to it.
What is clear is that m6A has fundamental roles in cell differentiation. Cells that lack the mark get stuck in a stem- or progenitor-like state. That can be lethal: when He and his colleagues disabled the m6A writer in mice, many embryos died in utero.
He has a possible explanation for the role of m6A. Each time a cell changes from one state to another — such as during differentiation — the mRNAs in it must change too. This change in mRNA content, which He calls a transcriptome switch, requires precision and careful timing. He thinks that the methyl marks might be a way for cells to synchronize the activity of thousands of transcripts.
As some researchers dive deep to try to understand the function of m6A and 6mA, others are looking for new modifications. Last year, He, Rechavi and their colleagues reported14 the discovery of another methyl mark on adenine in RNA called N1-methyladenosine (m1A). This mark also seems to promote translation, although the underlying mechanism is different from that of 6mA. He says it might also have a role in synchronizing transcripts for the transcriptome switch.
Then, in January, Jaffrey and his colleagues reported on yet another kind of modification that occurs near the caps of mRNAs. The researchers found that mRNAs with this mark — called m6Am — are more stable because their caps are harder to remove

Brian Wang is a Futurist Thought Leader and a popular Science blogger with 1 million readers per month. His blog Nextbigfuture.com is ranked #1 Science News Blog. It covers many disruptive technology and trends including Space, Robotics, Artificial Intelligence, Medicine, Anti-aging Biotechnology, and Nanotechnology.
Known for identifying cutting edge technologies, he is currently a Co-Founder of a startup and fundraiser for high potential early-stage companies. He is the Head of Research for Allocations for deep technology investments and an Angel Investor at Space Angels.
A frequent speaker at corporations, he has been a TEDx speaker, a Singularity University speaker and guest at numerous interviews for radio and podcasts. He is open to public speaking and advising engagements.