

In May, 2010, Venter and his team successfully inserted a fully customized strand of DNA into a living cell, creating what they call the “first synthetic genome.” Church says MAGE (Multiplex Automated Genome Engineering) can achieve similar results faster and cheaper. His lab’s device will go on sale later this year for about $90,000, and at least a dozen companies, including chemical giant DuPont (DD) and biotech startup Amyris, are considering purchasing it.
The new term is recombineering.
Recombineering (recombination-mediated genetic engineering)is a genetic and molecular biology technique based on homologous recombination systems, as opposed to the older/more common method of using restriction enzymes and ligases to cut and glue DNA sequences. It has been developed in E. coli and now is expanding to other bacteria species and is used to modify DNA in a precise and simple manner. The procedure is widely used for bacterial genetics, in the generation of target vectors for making a conditional mouse knockout, and for modifying DNA of any source often contained on a bacterial artificial chromosome (BAC).
New Scientist had updated coverage
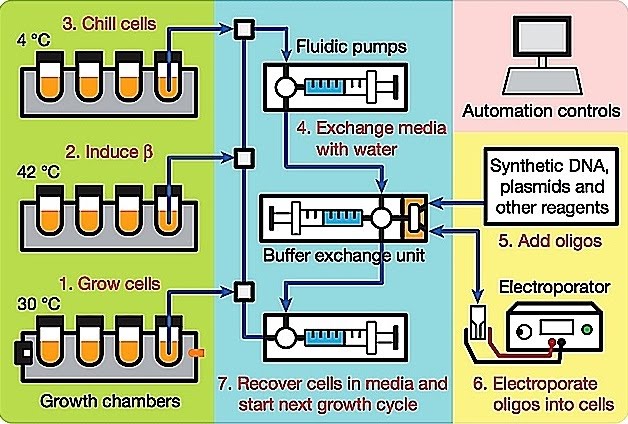
The machine let the E. coli multiply, mixed them with the DNA strands, and applied an electric shock to open up the bacterial cells and let the DNA get inside. There, some of the added DNA was swapped with the matching target sequences in the cells’ genomes. This process, called homologous recombination, is usually very rare, which is where the viral enzymes come in. They trick cells into treating the added DNA as its own, greatly increasing the chance of homologous recombination.
* Church is adapting MAGE for genetically modifying human stem cell lines. The work, funded by the US National Human Genome Research Institute, aims to create human cell lines with subtly different genomes in order to test ideas about which mutations cause disease and how.
* if MAGE really can be used to edit the genome of human cells, it would provide a way to fix the mutations that cause inherited disease. It could be the technology that opens the door to the genetic engineering of humans.
Recombineering utilizes linear DNA substrates that are either double-stranded (dsDNA) or single-stranded (ssDNA). Most commonly, dsDNA recombineering has been used to create gene replacements, deletions, insertions, inversions. Gene cloning and gene/protein tagging is also common. For gene replacements or deletions, usually a cassette encoding a drug-resistance gene is made by PCR using bi-partite primers. These primers consist of (from 5’→3’) 50 bases of homology to the target region, where the cassette is to be inserted, followed by 20 bases to prime the drug resistant cassette. The exact junction sequence of the final construct is determined by primer design. These events typically occur at a frequency of approximately 10^4/10^8cells that survive electroporation. Electroporation is the method used to transform the linear substrate into the recombining cell.
Recombineering with ssDNA provided a breakthrough both in the efficiency of the reaction and the ease of making point mutations. This technique was further enhanced by the discovery that by avoiding the methyl-directed mismatch repair system, the frequency of obtaining recombinants can be increased to over 10^7/10^8 viable cells. This frequency is high enough that alterations can now be made without selection. With optimized protocols, over 50% of the cells that survive electroporation contain the desired change. Recombineering with ssDNA only requires the Red Beta protein; Exo, Gamma and the host recombination proteins are not required. As proteins homologous to Beta and RecT are found in many bacteria and bacteriophages (over 100 as of February 2010), recombineering is likely to work in many different bacteria. Thus, recombineering with ssDNA is expanding the genetic tools available for research in a variety of organisms. To date, recombineering has been performed in E. coli, S. enterica, Y. pseudotuberculosis, and M. tuberculosis
The biggest advantage of recombineering is that it obviates the need for conveniently positioned restriction sites, whereas in conventional genetic engineering, DNA modification is often compromised by the availability of unique restriction sites. In engineering large constructs of over 100 kb, such as the Bacterial Artificial Chromosomes (BACs), or chromosomes, recombineering has become a necessity. Recombineering can generate the desired modifications without leaving any ‘footprints’ behind. It also forgoes multiple cloning stages for generating intermediate vectors and therefore is used to modify DNA constructs in a relatively short time-frame. The homology required is short enough that it can be generated in synthetic oligonucleotides and recombination with short oligonucleotides themselves is incredibly efficient. Recently, recombineering has been developed for high throughput DNA engineering applications termed ‘recombineering pipelines’. Recombineering pipelines support the large scale production of BAC transgenes and gene targeting constructs for functional genomics programs such as EUCOMM (European Conditional Mouse Mutagenesis Consortium) and KOMP (Knock-Out Mouse Program). Recombineering has also been automated, a process called “MAGE” -Multiplex Automated Genome Engineering, in the Church lab
Human MAGE
Next-Gen Reading and Writing of Microbial and Human Genomes talk by George Church
The cost of sequencing has plummeted a million fold in six years and integration with next-gen genome engineering is following a similar path. Our SynBERC SynBioSIS – BIOFAB group supports the community via high-throughput production and characterization of synthetic genes and genomes by novel and cost-effective resources like oligonucleotide chips and Multiplex Automated Genome Engineering (MAGE). Starting with up to 244K 300-mers or 1M 75-mers per chip (for as little as $500 per chip), with enzymatic error correction these assemble in 600-mers with error rates as good as 1/6000 (and with sequencing even better). E.coli MAGE (via ss-oligomers) can incorporate up to 5 mutations per 90-mer and up to seven 90-mers per cell per 2 hr. One MAGE device can produce up to 4 billion combinatorial genomes (cells) per day per each of 8 growth chambers. The engineered cells are characterized by FACS, automated microscopy, selective growth and quantitative sequencing assays of RNA and protein-NA interactions. Applications include optimization of metabolite, fuel, drug, and macromolecular production levels. New translation codes are aimed at efficient incorporation of multiple non-standard amino acids, multi-virus resistance and safety through nutritional and genetic isolation. For human MAGE we are optimizing various combinations of ss-oligos, Zn-finger and TALE targeting, ds-nucleases, deaminases, and recombinases. The SynBioSIS chip pipeline and E.coli MAGE help with constructing and selecting new ZnF and TALE and BACs for use in fibroblast, stem cells (hiPS) etc. Next-gen sequencing measures off-target mutational and epigenomic impacts and ratios of bar-codes.
If you liked this article, please give it a quick review on ycombinator or StumbleUpon. Thanks

Brian Wang is a Futurist Thought Leader and a popular Science blogger with 1 million readers per month. His blog Nextbigfuture.com is ranked #1 Science News Blog. It covers many disruptive technology and trends including Space, Robotics, Artificial Intelligence, Medicine, Anti-aging Biotechnology, and Nanotechnology.
Known for identifying cutting edge technologies, he is currently a Co-Founder of a startup and fundraiser for high potential early-stage companies. He is the Head of Research for Allocations for deep technology investments and an Angel Investor at Space Angels.
A frequent speaker at corporations, he has been a TEDx speaker, a Singularity University speaker and guest at numerous interviews for radio and podcasts. He is open to public speaking and advising engagements.